Linear systems of saddle-point type arise in a range of applications including optimization, mixed finite-element methods [1] for mechanics and fluid dynamics, economics, and finance. Due to their indefiniteness and generally unfavorable spectral properties, such systems are difficult to solve, particularly when their dimension is very large. In some applications - for example, when simulating fluid flow over large periods of time - such systems have to be solved many times over the course of a single run, and the linear solver rapidly becomes a major bottleneck. For this reason, finding an efficient and scalable solver is of the utmost importance.
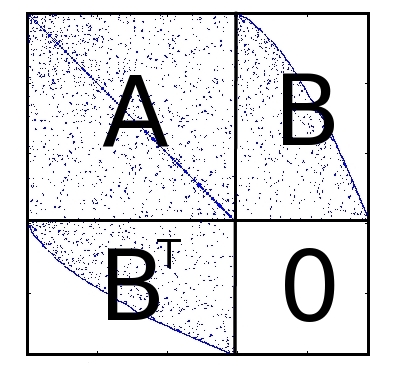
In this project, participants will be asked to propose and examine various solution strategies for saddle-point systems (see [2] for a very good, if slightly dated, survey). They will test the performance of those strategies on simple systems modeling flows in porous media. The different strategies will then be ranked based on their applicability, efficiency, and robustness.
Some knowledge of linear algebra and the basics of iterative solvers is expected. Familiarity with MATLAB is necessary.
References
[1] F. Brezzi and M. Fortin, Mixed and hybrid finite element methods, New York, Springer-Verlag, 1991.
[2] M. Benzi, G. H. Golub, J. Liesen, Numerical solution of saddle point problems, Acta Numerica (14), pp. 1-137, Cambridge University Press, 2005.


 .
.